Article Text

Statistics from Altmetric.com
Although considered by many in the west to be a rare and exotic infection, Japanese encephalitis is numerically one of the most important causes of viral encephalitis worldwide, with an estimated 50 000 cases and 15 000 deaths annually.1 2 About one third of patients die, and half of the survivors have severe neuropshychiatric sequelae. Most of China, Southeast Asia, and the Indian subcontinent are affected by the virus, which is spreading at an alarming rate. In these areas, wards full of children and young adults afflicted by Japanese encephalitis attest to its importance.
Historical perspective
Epidemics of encephalitis were described in Japan from the 1870s onwards. Major epidemics were reported about every 10 years, with more than 6000 cases reported in the 1924 epidemic.3 The term type B encephalitis was originally used to distinguish these summer epidemics from von Economo's encephalitis lethargica (sleeping sickness, known as type A), but the B has since been dropped. In 1933 a filterable agent was transmitted from the brain of a fatal case to cause encephalitis in monkeys; the prototype Nakayama strain of Japanese encephalitis virus was isolated from the brain of a fatal case in 1935. The virus was later classed as a member of the genus Flavivirus (family Flaviviridae) named after the prototype yellow fever virus (Latin; yellow=flavi). Although of no taxonomic significance, the ecological term arbovirus is often used to describe the fact that Japanese encephalitis virus is insect (arthropod) borne.
Epidemiology
NEUROTROPIC FLAVIVIRUSES: A GLOBAL PERSPECTIVE
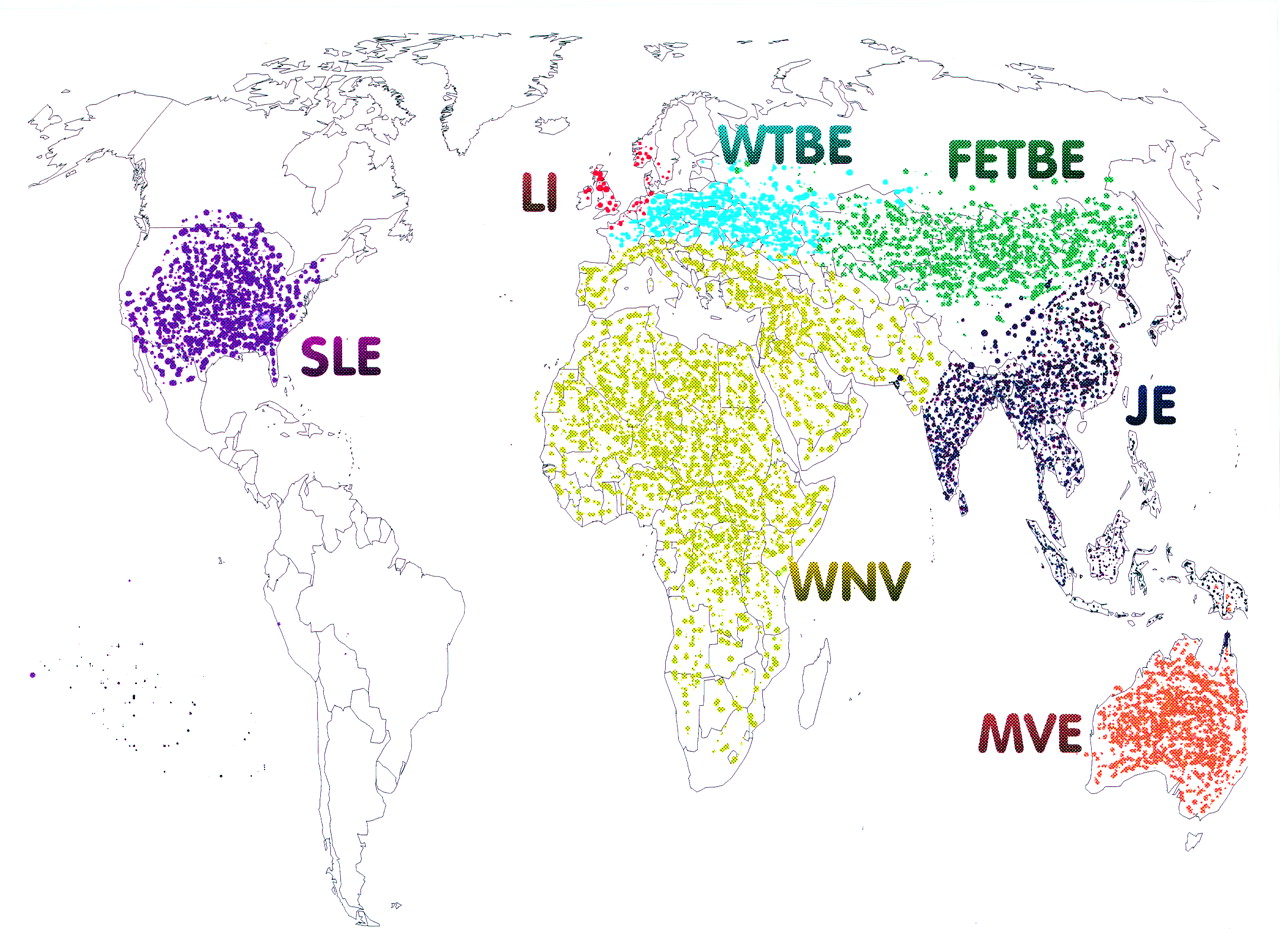
Japanese encephalitis virus is transmitted between animals by Culex mosquitoes, and occurs across eastern and southern Asia and the Pacific rim. However, related neurotropic flaviviruses are found across the globe (fig 1); they share many virological, epidemiological, and clinical features.2 Molecular virological studies suggest that all flaviviruses derived from a common ancestor some 10–20 000 years ago, and are rapidly evolving to fill ecological niches.4 Examples of mosquito borne neurotropic flaviviruses include Murray Valley encephalitis virus in Australia, and St Louis encephalitis virus in North America. West Nile virus, a flavivirus found in Africa, the Middle East, and parts of Europe, is traditionally associated with a syndrome of fever arthralgia and rash, and with occasional nervous system disease. However, in 1996 West Nile virus caused an outbreak of encephalitis in Romania,5 and a West Nile-like flavivirus was responsible for an encephalitis outbreak in New York in 1999.6 7

Map showing approximate global distribution of major neurotropic flaviviruses; JE=Japanese encephalitis; MVE=Murray valley encephalitis; WN=West Nile; WTBE=Western tick-borne encephalitis; FETBE=Far Eastern tick-borne encephalitis; LI=Louping Ill virus; SLE=St Louis encephalitis.
In northern Europe and northern Asia, flaviviruses have evolved to use ticks as vectors because they are more abundant than mosquitoes in cooler climates. Far eastern tick-borne encephalitis virus (also known as Russian spring-summer encephalitis virus) is endemic in the eastern part of the former USSR, and Western tick-borne encephalitis virus occurs in Europe and has caused recent epidemics in Germany and Austria.8 In the United Kingdom the tick borne Louping Ill virus is is enzootic in sheep, and occasionally causes encephalitis in sheep and humans.9
ENZOOTIC CYCLE
Japanese encephalitis virus is transmitted naturally between wild and domestic birds, and pigs by Culex mosquitoes—the most important for human infection being Culex tritaeniorrhynchus which breeds in pools of stagnant water (such as rice paddy fields).10 Although many animals can be infected with the virus, only those which develop high viraemias are important in the natural cycle. As well as maintaining and amplifying Japanese encephalitis virus in the environment, birds may also be responsible for the spread to new geographical areas. Pigs are the most important natural host for transmission to humans, because they are often kept close to humans, have prolonged and high viraemias, and produce many offspring—thus providing a continuous supply of previously uninfected new hosts. The virus does not typically cause encephalitis in these natural hosts, athough abortions occur in pregnant sows.
EPIDEMIOLOGY OF HUMAN DISEASE
Humans become infected with Japanese encephalitis virus coincidentally when living or travelling in close proximity to the enzootic cycle of the virus. Although most cases occur in rural areas, Japanese encephalitis virus is also found on the edge of cities. Epidemiological studies have shown that after the monsoon rains mosquitoes breed prolifically, and as their numbers grow, so does their carriage of Japanese encephalitis virus and the infection rate of pigs.11 12 Human infection soon follows. In sentinel studies, previously unexposed pigs placed in endemic areas were infected with the virus within weeks.
Although the virus has occasionally been isolated from human peripheral blood13 viraemias are usually brief and titres low; thus humans are considered a dead end host from which transmission does not normally occur. Cross sectional serological surveys have shown that in rural Asia most of the population are infected with Japanese encephalitis virus during childhood or early adulthood.14About 10% of the susceptible population is infected each year.15 However, most infections of humans are asymptomatic or result in a non-specific flu-like illness; estimates of the ratio of symptomatic to asymptomatic infection vary between 1 in 2516 and 1 in 1000.17
Japanese encephalitis is mostly a disease of children and young adults. In northern Thailand the incidence has been estimated to be up to 40 per 100 000 for ages 5 to 25, declining to almost zero for those over 3514 18 The incidence is lower among young children (<3 years old) than in older children, possibly reflecting behavioural factors—for example, playing outside after dusk.19
When epidemics first occur in new locations, such as in Sri Lanka, India, and Nepal, adults are also affected.20 The susceptibility of immunologically naive adults was also demonstrated by the incidence of Japanese encephalitis among American troops during conflicts in Japan, Korea, and Vietnam21-25 The rate of symptomatic infection was higher in these troops than in local populations. This may be explained by partial protection due to previous flavivirus exposure in the indigenous population, age related differences, different genetic susceptibility to Japanese encephalitis, or more sensitive disease surveillance among United States troops.
Broadly speaking two epidemiological patterns of Japanese encephalitis are recognised.19 In northern areas (northern Vietnam, northern Thailand, Korea, Japan, Taiwan, China, Nepal, and northern India) huge epidemics occur during the summer months, whereas in southern areas (southern Vietnam, southern Thailand, Indonesia, Malaysia, Philippines, Sri Lanka, and southern India) Japanese encephalitis tends to be endemic, and cases occur sporadically throughout the year with a peak after the start of the rainy season.19
Various explanations for this different pattern have been offered. The finding that Japanese encephalitis virus isolates from epidemic northern Thailand and endemic southern Thailand were of different genotypes (see below) led to the suggestion that differing neurovirulence among different strains may be responsible.26 However, data from Vietnam do not support this: isolates of the virus from epidemic northern Vietnam were the same genotype as those from endemic southern Vietnam.27Comparisons of climatic data from northern and southern Vietnam suggest that temperature may be important (fig 2). Whereas rainfall patterns are almost identical in northern and southern Vietnam, the temperature is very different, and the number of cases of encephalitis seems to follow temperature closely. In the south, where the temperature remains high through the year, the number of cases each month is constant. In the north a sharp rise in cases of Japanese encephalitis during the summer months corresponds with a rise in temperature above 20°C. The prolonged mosquito larval development time and longer extrinsic incubation period of Japanese encephalitis virus at cooler temperature, which thus reduce the rate of virus transmission, could be one explanation for these findings.
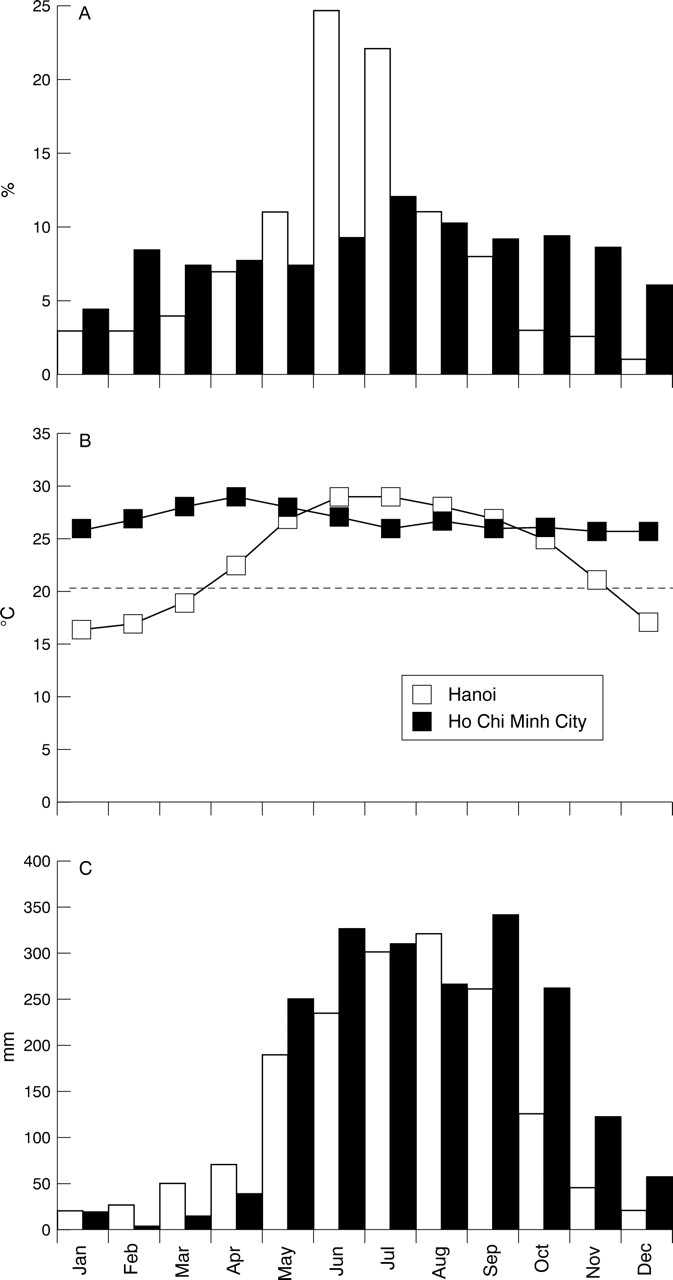
Relation between (A) encephalitis cases, (B) temperature, and (C) rainfall in Ho Chi Minh city (southern Vietnam), and Hanoi (northern Vietnam). (Source: National Institute of Health and Epidemiology, Hanoi, and Pasteur Institute, Ho Chi Minh City.)
GEOGRAPHICAL DISTRIBUTION
In the past 50 years the geographical area affected by Japanese encephalitis virus has expanded (fig 1). Differences in diagnostic capabilities and in reporting of encephalitis make it impossible to plot this expansion precisely. However, the timing of the first reported cases or new epidemics in each area gives an impression of the relentless spread of Japanese encephalitis. In China outbreaks of summer encephalitis occurred from 1935, and the virus was first isolated there in 1940; there are currently 10–20 000 cases a year, although in the early 1970s it was over 80 000 cases annually.19 In the far eastern Russian states, Japanese encephalitis first occurred in 1938. In 1949, large epidemics were reported from South Korea for the first time. Epidemics in northern Vietnam followed in 1965 (currently 1000–3000 cases nationally a year), and in Chiang Mai in northern Thailand in 1969 (currently 1500–2500 cases nationally a year). Japanese encephalitis was recognised in southern India from 1955, but was confined to the south until the 1970s. Since then, large outbreaks (2000–7000 cases a year) have been reported from eastern and northeastern states. The fact that adults and children were equally affected in these Indian states strongly supports the idea that the virus was introduced here for the first time. The late 1970s also saw the first cases in Burma and Bangladesh, and large epidemics (up to 500 cases a year) in southwestern Nepal. In 1985 Sri Lanka experienced its first epidemic with 410 cases and 75 deaths. Japanese encephalitis virus continues to spread west with cases occurring in Pakistan28 and new epidemics in the Kathmandu valley of Nepal.29
Charting the progression of the disease southeast across Asia and the Pacific rim is harder because sporadic cases in endemic areas do not command the same attention as the massive epidemics that occur in temperate climates. The disease has occurred on the western Pacific islands with outbreaks in Guam in 194730 and Saipan in 1990.31 In Malaysia the disease is endemic; the virus was first isolated in the 1960s and about 100 cases are recorded annually. The epidemiology has recently been complicated by a superimposed epidemic of a previously unidentified encephalitic virus. This RNA paramyxovirus (named Nipah virus) is similar to the Australian Hendra virus and seems to be transmitted to humans (especially abattoir workers and farmers) from the bodily fluid of pigs.32-34Japanese encephalitis is endemic in Indonesia, and 1000–2500 cases of encephalitis are reported annually, although in most the aetiological agent is not confirmed.35 Further east, Japanese encephalitis occurs sporadically in the Philippines and New Guinea. The first cases occurred in the Australian Torres Straits islands in 1995,36 and it was reported for the first time north of Cairns on the Australian mainland in 1998.37 38
The reasons for the spread of Japanese encephalitis are incompletely understood, but probably include changing agricultural practices, such as increasing irrigation (which allows mosquito breeding), and animal husbandry (which provides host animals). In Indonesia, the lower prevalence of antibody to Japanese encephalitis virus in Borneo than neighbouring Bali has been attributed to the lack of pigs in this predominantly Moslem culture.35 In developed countries such as Japan, Taiwan, and South Korea the number of cases has fallen, probably due to a combination of mass vaccination of children, spraying of pesticides, changing pig rearing practices, separation of housing from farming, better housing with air conditioning, and less availability of mosquito breeding pools.10 However, in Korea the widespread use of vaccine in children has been associated with a higher incidence of Japanese encephalitis in those over 15 years.19
Virology
In common with all flaviviruses, Japanese encephalitis virus has a small (50 nm) lipoprotein envelope surrounding a nucleocapsid comprising of core protein and 11 kb single stranded RNA (3800 kD). At least five genotypes of Japanese encephalitis virus occur in Asia, which relate roughly to the geographical area of isolation.26 39 The complete nucleotide sequence has been published, and includes 5′ and 3′ untranslated regions, and a single open reading frame encoding genes for three structural proteins (capsid protein (C); precursor to the membrane M protein (PrM); and envelope protein (E)) and seven non-structural proteins. The search for genetic determinants of virulence in animal models of flavivirus encephalitis has focused on the E protein.40 This protein, of about 500 amino acids, is the major component of the surface projections of the virion. As well as eliciting neutralising antibodies and protective immune responses in the host41 42 it is thought to be the cell receptor binding protein and mediator of membrane fusion and cell entry.43 A highly sulphated heparan sulphate molecule has recently been identified as the putative receptor of flavivirus cell entry.44 Various approaches have allowed E gene sequences of flaviviruses to be related to virulence in animal models. These suggest that the E protein has a major role in determination of virulence phenotype, and that single amino acid substitutions are sufficient to cause loss of neurovirulence or neuroinvasiveness.45-47 Whether such differences are important in determining the clinical presentation of Japanese encephalitis virus in humans is unknown.
Clinical features
Patients with Japanese encephalitis typically present after a few days of non-specific febrile illness, which may include coryza, diarrhoea, and rigors.2 This is followed by headache, vomiting, and a reduced level of consciousness, often heralded by a convulsion. In some patients, particularly older children and adults, abnormal behaviour may be the only presenting feature, resulting in an initial diagnosis of mental illness. For example, during the Korean conflict some American servicemen with Japanese encephalitis were initially diagnosed as having “war neurosis”.23
A proportion of patients make a rapid spontaneous recovery (so called abortive encephalitis). Others may present with aseptic meningitis and have no encephalopathic features.48Convulsions occur often in Japanese encephalitis, and have been reported in up to 85% of children49 and 10% of adults.22 50 In some children a single convulsion is followed by a rapid recovery of consciousness, resulting in a clinical diagnosis of febrile convulsion. Generalised tonic-clonic seizures occur more often than focal motor seizures. Multiple or prolonged seizures and status epilepticus are associated with a poor outcome (Solomon T et al, unpublished observations). In a proportion of children subtle motor seizures occur, causing twitching of a digit, eye, or mouth, eye deviation, nystagmus, excess salivation, or irregular respiration. Without electroencephalographic monitoring these may be difficult to identify (Solomon Tet al, unpublished observations).
The classic description of Japanese encephalitis includes a dull flat mask-like facies with wide unblinking eyes (fig 3), tremor, generalised hypertonia, and cogwheel rigidity. These features were reported in 70%-80% of American service personnel, and 20%-40% of Indian children.48 49 Opisthotonus and rigidity spasms, particularly on stimulation, occur in about 15% of patients and are associated with a poor prognosis.48 49 Other extrapyramidal features include head nodding and pill rolling movements, opsoclonus myoclonus, choreoathetosis, and bizarre facial grimacing, and lip smacking (fig 4).48 49 51 Upper motor neuron facial nerve palsies occur in around 10% of children and may be subtle, or intermittent.

Staring mask-like facies due to a wide palpebral angle in two Vietnamese children with Japanese encephalitis. (T Solomon.)
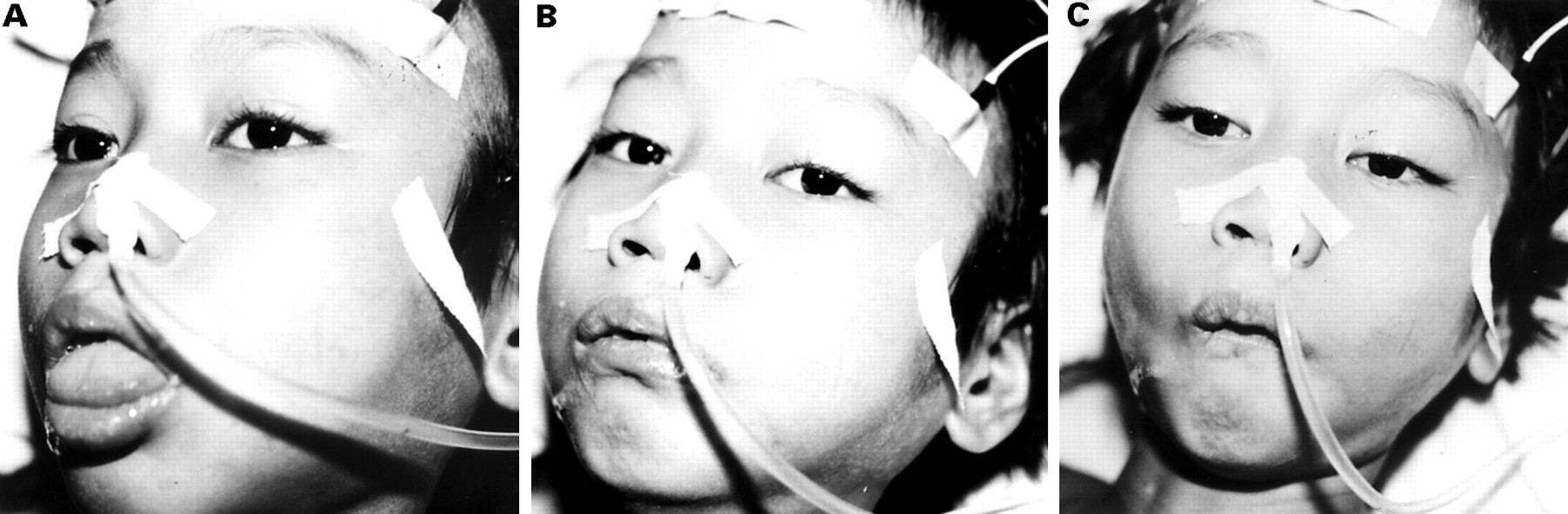
Facial grimacing in a Vietnamese boy with Japanese encephalitis. (T Solomon.)
Changes of respiratory pattern, flexor and extensor posturing, and abnormalities of the pupillary and occulocephalic reflexes are poor prognostic signs.48 49 52 and may reflect encephalitis in the brain stem.10 However in some patients a clear rostrocaudal progression of brainstem signs, an association with high CSF opening pressures, and a reversal of signs on aggressive management of raised intracranial pressure suggests that transtentorial herniation may also contribute.(Solomon T et al, unpublished observations)
ACUTE FLACCID PARALYSIS
Recently we have identified a subgroup of patients infected with Japanese encephalitis virus who presented with a poliomyelitis-like acute flaccid paralysis.53 After a short febrile illness there was a rapid onset of flaccid paralysis in one or more limbs, despite a normal level of consciousness. Weakness occurred more often in the legs than the arms, and was usually asymmetric. Thirty per cent of such patients subsequently developed encephalitis, with reduced level of consciousness, and upper motor neuron signs, but in most acute flaccid paralysis was the only feature. At follow up (1–2 years later) there was persistent weakness and marked wasting in the affected limbs. Nerve conduction studies demonstrated markedly reduced motor amplitudes, and EMG showed a chronic partial denervation, suggesting anterior horn cell damage.53 Flaccid paralysis also occurs in comatose patients with “classic” Japanese encephalitis, being reported in 5%-20%.22 54 Electrophysiological studies have confirmed anterior horn cell damage, and MRI of the spinal cord showed abnormal signal intensity on T2 weighted images.55Occasionally respiratory muscle paralysis may be the presenting feature.56
OUTCOME
About 30% of patients admitted to hospital with Japanese encephalitis die, and around half of the survivors have severe neurological sequelae. However, in areas with better hospital facilities there is a reduction in mortality, but a concomitant increase in the number of patients with sequelae.2 About 30% of survivors have frank motor deficits. These include a mixture of upper and lower motor neuron weakness, and cerebellar and extrapyramidal signs.24 57 Fixed flexion deformities of the arms, and hyperextension of the legs with “equine feet” are common (fig 5). Twenty per cent of patients have severe cognitive and language impairment (most with motor impairment also), and 20% have further convulsions.58 59 A higher rate of sequelae is reported for children than adults.60 In addition, more detailed studies have shown that about half of those who were classed in the good recovery group have more subtle sequelae such as learning difficulties, behavioural problems, and subtle neurological signs.58

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Fixed flexion of the upper limb: a common sequelae in Japanese encephalitis.
INVESTIGATIONS
A peripheral neutrophil leukocytosis is seen in most patients, and hyponatraemia may occur as a consequence of inappropriate antidiuretic hormone secretion (SIADH). The CSF opening pressure is increased in about 50% of patients. High pressures (>250 mm) are associated with a poor outcome (Solomon T et al, unpublished observations). Typically there is a moderate CSF pleocytosis of 10–100 cells/mm3, with predominant lymphocytes, mildly increased protein (50–200 mg%), and a normal glucose ratio. However, polymorphonuclear cells may predominate early in the disease, or there may be no CSF pleocytosis.49
In about 50% of patients CT shows bilateral non-enhancing low density areas in one or more of the thalamus, basal ganglia, midbrain, pons, and medulla.61 62 Magnetic resonance imaging is more sensitive, typically demonstrating more extensive lesions, (typically high signal intensity on T2 weighted images) of the thalamus, cerebral hemispheres, and cerebellum.55 63 Thalamic lesions of mixed intensity may also be seen on T1 and T2 weighted scans suggesting haemorrhage.51 55 Imaging studies may be useful in distinguishing Japanese encephalitis from herpes simplex encephalitis, where the changes are characteristically frontotemporal.64However, most reports are of scans performed late in the illness, and the diagnostic value of scans performed early is unknown. Single photon emission tomography (SPECT) studies carried out acutely may show hyperperfusion in the thalamus and putamen.65 Follow up studies have shown hypoperfusion in the same areas, as well as in the frontal lobes.51
Various electroencephalographic abnormalities have been reported in Japanese encephalitis including theta and delta coma, burst suppression, epileptiform activity, and occasionally alpha coma.62 66 Diffuse slowing may be useful in distinguishing Japanese encephalitis from herpes simplex virus, in which changes are characteristically frontotemporal.64Measurement of evoked potentials shows delays of central motor conduction times consistent with widespread involvement at the cortical and subcortical levels.62
The differential diagnosis of Japanese encephalitis is broad and includes other viral encephalitides (arboviruses, herpes viruses, enteroviruses, and postinfectious and postvaccination encephalomyelitis), other CNS infections (bacterial and fungal meningitis, tuberculosis, cerebral malaria, leptospirosis, tetanus, abscesses), other infectious diseases with CNS manifestations (typhoid encephalopathy, febrile convulsions), and non-infectious diseases (tumours, cerebrovascular accidents, Reye's syndrome, toxic and alcoholic encephalopathies, and epilepsy).2 10 Where other flaviviruses circulate they should also be included in the differential. Even viruses which are not traditionally considered as neurotropic may cause CNS disease, and only with appropriate diagnostic tests can viruses such as West Nile virus and dengue viruses be distinguished from Japanese encephalitis virus.67 67a
DIAGNOSIS
Attempts to isolate Japanese encephalitis virus from clinical specimens are usually unsuccessful, probably because of low viral titres and the rapid production of neutralising antibodies. Isolates may sometimes be obtained from CSF (in which case it is associated with a failure of antibody production and a high mortality rate)68 or from brain tissue (either at necropsy or postmortem needle biopsy). Immunohistochemical staining of CSF cells or necropsy tissue with anti- Japanese encephalitis virus polyclonal antibodies may be positive.69 70 However, for most practical purposes Japanese encephalitis is diagnosed serologically. The haemaglutination inhibition test was used for many years, but it had various practical limitations, and as it required paired serum, could not give an early diagnosis.71 In the 1980s IgM and IgG capture enzyme linked immunosorbant assays (ELISAs) were developed which have become the accepted standard for diagnosis of Japanese encephalitis.72 73 After the first few days of illness, the presence of anti-Japanese encephalitis virus IgM in the CSF has a sensitivity and specificity of >95% for CNS infection with the virus (before this false negatives may occur).74 However, because ELISAs require complex equipment, their use has been confined largely to a few academic or referral centres, rather than the rural areas where Japanese encephalitis occurs. Recently the IgM ELISA has been modified to a simple nitrocellulose membrane based format in which the result is a colour change visible to the naked eye.75This test, which is rapid, simple to use, and requires no specialised equipment should prove useful for diagnosis of the disease in rural hospitals. Japanese encephalitis virus RNA has been detected in human CSF samples using the reverse transcriptase polymerase chain reaction28 76; however, its reliability as a routine diagnostic test has yet to be shown.
Pathogenesis
Only about 1 in 25 to 1 in 1000 humans infected with Japanese encephalitis virus develop clinical features of infection.16 17 19 These may range from a mild flu-like illness to a fatal meningoencephalomyelitis. The factors determining which of all the humans infected develop disease are unknown, but could include viral factors such as route of entry, titre, and neurovirulence of the inoculum, and host factors such as age, genetic make up, general health, and pre-existing immunity.
After the bite of an infected mosquito, the virus is thought to amplify peripherally, causing a transient viraemia before invading the CNS. Based on data from mice and macaque monkeys, the site of peripheral amplification is thought to be dermal tissue and then lymph nodes. The means by which Japanese encephalitis virus crosses the blood-brain barrier is unknown. In experimental studies with a hamster model of St Louis encephalitis virus (a related flavivirus) the olfactory route was shown to be important.77 Intranasal spraying is also an effective means of experimentally inoculating monkeys.78However, immunohistochemical staining of human postmortem material has shown diffuse infection throughout the brain, indicating a haematogenous route of entry.70 79 Although experimental evidence suggests that replication within endothelial cells may be an important means of crossing the blood-brain barrier in some flaviviruses, for Japanese encephalitis virus passive transfer across the endothelial cells seems a more likely mechanism.80 81Other factors which compromise the integrity of the blood-brain barrier have also been implicated as risk factors for neuroinvasion. In several studies a disproportionate number of fatal cases had neurocysticercosis at necropsy.82 83 It has also been suggested that head trauma (for example, due to a road traffic accident) during the transient viraemia could facilitate viral entry into the CNS.84
Electron microscopic studies of the brains of infected mice show that the virus replicates in the rough endoplasmic reticulum and golgi apparatus. There is hypertrophy of the endoplasmic reticulum and degeneration into cyctic structures causing extensive dysfunction.85
HISTOPATHOLOGY
At necropsy, CNS findings in Japanese encephalitis reflect the inflammatory response to widespread neuronal infection with virus.3 79 86 The leptomeninges are normal or hazy. The brain parenchyma is congested with focal petechiae or haemorrhage in the grey matter. When survival is prolonged beyond 7 days blotchy necrolytic zones are seen. The white matter usually appears normal. In some patients, the grey matter of the spinal cord is confluently discoloured, resembling that of poliomyelitis.87 The thalamus, basal ganglia, and midbrain are heavily affected, providing anatomical correlates for the tremor and dystonias which characterise Japanese encephalitis. At the histological level, invasion of neurons by Japanese encephalitis virus is followed by perivascular cuffing, infiltration of inflammatory cells (T cells and macrophages) into the parenchyma, and phagocytosis of infected cells.3 79 T cells in the brains of fatal cases stained with monoclonal antibodies are CD8+ and CD8- (presumed to be CD4+) and are localised at the perivascular cuff. Both cell types are found in the CSF in acute infection, though the predominant cell type is CD4+.79 In patients that die rapidly, there may be no histological signs of inflammation, but immunohistochemical studies disclose viral antigen in morphologically normal neurons.79 88 This may explain the normal CSF findings in a proportion of patients with Japanese encephalitis.
IMMUNOLOGY
Interferon and interferon inducers are active against Japanese encephalitis virus in mice and monkeys,89 90 and endogenous interferon-α has been detected in the plasma and CSF of humans with Japanese encephalitis.91 In addition both humoral and cellular immune responses occur after infection with Japanese encephalitis virus. The humoral immune response in Japanese encephalitis has been well characterised. When disease is due to primary infection (when Japanese encephalitis virus is the first flavivirus with which a person has been infected) a rapid and potent IgM response occurs in serum and CSF within days of infection. By day 7 most patients have raised titres.74 Attempts to isolate the virus are usually negative in such patients. However, the failure to mount an IgM response is associated with positive virus isolation and a fatal outcome.68 Antibody to Japanese encephalitis virus probably protects the host by restricting viral replication during the viraemic phase, before the virus crosses the blood-brain barrier.92 Evidence from other flaviviruses suggests that it may also limit damage during established encephalitis by neutralising extracellular virus and facilitating lysis of infected cells by antibody dependent cellular cytotoxicity.93
In surviving patients immunoglobulin class switching occurs, and within 30 days most have IgG in the serum and CSF. Asymptomatic infection with Japanese encephalitis virus is also associated with raised IgM in the serum, but not CSF.74 In patients with secondary infection (those who have previously been infected with a different flavivirus—for example, dengue infection or yellow fever vaccination) there is an anamnastic response to flavivirus group common antigens.74 This secondary pattern of antibody activation is characterised by an early rise in IgG with a subsequent slow rise in IgM.
CELLULAR IMMUNITY
In animal models of Japanese encephalitis, the cellular immune response seems to contribute to the prevention of disease during acute infection by restricting virus replication before the CNS is invaded: athymic nude mice have increased susceptibility to experimental infection with Japanese encephalitis virus94; transfer of spleen cells from mice immunised with live attenuated virus conveys immunity to infection.95 Spider monkeys, which are normally unaffected by intracerebrally inoculated virus develop rapidly progressive encephalitis when T cell function has been impaired by cyclophosphamide.96
In humans infected with St Louis encephalitis virus (a flavivirus in the same antigenic complex as Japanese encephalitis virus) impairment of T cell function by human immunodeficiency virus (HIV) seems to increase the risk of developing encephalitis.97 By analogy with other human viral infections, including influenza, HIV, Epstein-Barr virus, and dengue, cytotoxic T lymphocytes might be important in the control and possibly clearance of Japanese encephalitis virus.98 99 Preliminary experimental evidence is in agreement with this: T lymphocyte responses were characterised in seven convalescent patients with Japanese encephalitis and 10 vaccine recipients.100 Japanese encephalitis virus specific T cell proliferation (including CD4+ and CD8+ T lymphocyte responses) was demonstrated in both groups. Japanese encephalitis virus specific and flavivirus cross reactive CD4+ T lymphocytes which recognise E protein in an HLA restricted manner were recently demonstrated in two vaccine recipients.101
Management
Treatment for Japanese encephalitis is supportive, and involves controlling convulsions and raised intracranial pressure when they occur. For many years corticosteroids were given, but a double blind randomised placebo controlled trial of dexamethasone failed to show any benefit.52 Careful nursing care and physiotherapy are needed to reduce the risk of bed sores, malnutrition, and contractures. Aspiration pneumonia is a common occurrence in patients with a reduced gag reflex. There is currently no specific treatment for Japanese encephalitis. Isoquinolone compounds are effective in vitro,102 and monoclonal antibodies are apparently effective in animal models.103 104 Interferon-α is currently the most promising potential treatment. It is produced naturally in the CSF in response to infection with Japanese encephalitis virus91 and in vitro it has activity against the virus.105 Recombinant interferon-α has been given in open trials to a few patients with encouraging results,106and is currently being assessed in a placebo controlled double blind trial.
Prevention
Broadly speaking, measures to control Japanese encephalitis include those which interfere with the enzootic cycle of the virus, and those which prevent disease in humans. Measures to control breeding of Culex mosquitoes, such as the application of larvicides to rice fields, and insecticide spraying have proved ineffectual.10Inactivated and live attenuated vaccines (described below) have been used to protect swine against the virus; however, widespread vaccination is not feasible in most settings. Residents and travellers to endemic areas should take personal protection to reduce the number of Culex bites. These include minimising outdoor exposure at dusk and dawn, wearing clothing that leaves a minimum of exposed skin, using insect repellents containing at least 30% DEET (N,N-diethyl-3 methlybenzamide) and sleeping under bed nets. While these measures may be possible for the short term visitor, most are not practical for residents of endemic areas.
FORMALIN INACTIVATED VACCINE
Formalin inactivated vaccines against Japanese encephalitis were produced in Russia, Japan, and in the United States by Albert Sabin (later of polio fame) during the second world war to protect American troops in Asia.107 A similar formalin inactivated vaccine has been manufactured in Japan since 1954. It is produced by Osaka University and is available internationally under the BIKEN label. Similar vaccines are made by other manufacturers in India, Japan, Korea, Taiwan, Thailand, and Vietnam. The vaccine's efficacy was shown in large double blind randomised tetanus toxoid controlled trials in Taiwan and Thailand involving more than 300 000 children.14
In western subjects three doses of vaccine are required to give protective antibody levels to a suitably high number of recipients (80%-100%); It is given at 0, 7, and 30 days, with a booster immunisation recommended at 1 year. In Asian subjects two doses may be sufficient because of prior or subsequent exposure to Japanese encephalitis or other flaviviruses (for example, West Nile virus, dengue virus). A booster vaccination has been recommended at 1–2 years for those with continued exposure; however, a recent follow up study of a cohort of immunised adults showed persistence of antibody at 3 years after the initial course.108
ADVERSE REACTIONS
Japanese encephalitis vaccination is associated with a moderate frequency of local and mild systemic side effects. Tenderness, redness, and swelling has been reported in up to 20% of vaccine recipients, and fever, headache, malaise, and chills have been reported in about 10%. Because the vaccine is derived from mouse brain there has been concern about neurologically related side effects. However, the amount of mouse myelin basic protein in the vaccine is negligible. Surveillance of United States vaccine recipients in 1945 showed no increase of neurological conditions above background levels, and there were no neurological events among 44 000 Thai children receiving vaccine in an efficacy trial.14 In Japan, surveillance of Japanese encephalitis vaccine related complications between 1965 and 1973 disclosed neurological events (encephalitis, encephalopathy, seizures, peripheral neuropathy) at a rate of 1–2.3/million recipients, which is less than reported for other virus vaccines. However, since 1989 a new pattern of adverse events has been recognised among European, American, and Australian vaccine recipients.109 These take the form of itching, urticaria, and occasionally angio-oedema of the face—sometimes requiring admission to hospital and corticosteroid therapy. Many of these reactions occurred several days after vaccination. The incidence of these reactions was estimated at 2 to 10 per 1000 vaccinees, and was more likely in those with a history of urticaria.110 No vaccine constituent responsible for this apparently new adverse reaction has been identified, although some cases have been associated with allergy to gelatin stabiliser.
WHO SHOULD BE VACCINATED?
Japanese encephalitis vaccine is recommended for native and expatriate residents of endemic areas, laboratory workers potentially exposed to the virus, and for travellers spending 30 days or more in endemic areas.111 For shorter visits, the vaccine is only recommended if there will be extensive outdoor activity in rural areas, or if visiting during known epidemics.111 However, considering the variable incidence of Japanese encephalitis from year to year, its unpredictability, and the unreliability of some epidemiological data, identifying areas of epidemic transmission is difficult. It has been argued that the benefit of immunisation exceeds the risk of vaccine related adverse effects, especially when the devastating impact of acquiring Japanese encephalitis is contrasted with the lesser risk of an allergic reaction that can be aborted by drug therapy.10 Two recent cases of the disease in short term (<2 weeks) visitors to Bali would support the contention that all travellers to endemic areas should be vaccinated.106-108
LIVE ATTENUATED VACCINE
In 1988 the Chinese authorities licensed a new live attenuated Japanese encephalitis vaccine. This strain (SA 14–14–2) was produced by passing the virus through weanling mice, then culturing in primary baby hamster kidney cells. The vaccine has been shown to be safe and immunogenic, and has been given to over 100 million children in China. Its efficacy was recently demonstrated in a relatively simple and inexpensive case-control study in which the prevalence of immunisation was compared between 56 cases of Japanese encephalitis and 1299 age and village matched controls.112 The effectiveness of one dose was 80% (95% confidence intervals 44%-93%) and of two doses 1 year apart 97.5% (86%-99.6%). The vaccine's short term safety was recently confirmed in a randomised trial of 26 000 children,113 and it has been shown to be immunogenic at the shorter dosage interval of 1 and 2.5 months, which might facilitate its incorporation into existing immunisation programmes.114
Future directions
Despite some successes with formalin inactivated vaccination, and the promise of the new live attenuated vaccine, Japanese encephalitis looks likely to remain an important public health problem into the next millennium. Unlike smallpox and polio, for which humans are the only host and elimination by vaccination is feasible, the enzootic nature of Japanese encephalitis virus means that there is no possibility of global eradication. The geographical area affected is expanding, and the 2.8 billion people living in affected areas will continue to be exposed to the virus. Thankfully, only a small proportion of them develops disease, but we have little understanding of what determines who develops disease and how it will manifest. The recent discovery of a poliomyelitis-like presentation of Japanese encephalitis virus in Vietnam raises important questions, especially as the target for global eradication of polio by the year 2000 approaches. Is this Japanese encephalitis virus myelitis unique to Vietnam, or is it also important in other areas? Has it recently arisen, or might patients who were previously labelled as having polio, actually have been infected with Japanese encephalitis virus? The viral and host factors which determine who develops disease and the differing clinical presentations need further investigation. Virus isolates of varying pathogenicity for mice occur, and small changes in the virus envelope protein affect neurovirulence. Whether such differences are important in humans needs to be investigated.
Considering the many cases of Japanese encephalitis, research into antiviral drugs has been relatively neglected. Interferon-α, which was shown to be effective in vitro and in animal models nearly 15 years ago is only now being assessed in human disease. Attention should focus on newer antiviral drugs, and their possible role in Japanese encephalitis. Pathophysiological research has suggested possible therapeutic avenues, even in the absence of specific antiviral drugs. Findings from Vietnam on the importance of seizures and raised intracranial pressure in the disease need to be confirmed in other settings, and intervention trials considered. Japanese encephalitis virus is expanding across the globe at an alarming rate. New rapid diagnostic methods should facilitate monitoring the spread of the disease in locations where, until now, the aetiology of encephalitis could only be guessed. The environmental and ecological factors responsible for this expansion need further investigations, with a view to controlling the spread of this fascinating and devastating disease.
Acknowledgments
We thank our many colleagues in Southeast Asia who have contributed to some of the work and ideas contained in this article. Some of the work described was supported by the Wellcome Trust of Great Britain.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 67a.
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.↵
- 92.↵
- 93.↵
- 94.↵
- 95.↵
- 96.↵
- 97.↵
- 98.↵
- 99.↵
- 100.↵
- 101.↵
- 102.↵
- 103.↵
- 104.↵
- 105.↵
- 106.↵
- 107.↵
- 108.↵
- 109.↵
- 110.↵
- 111.↵
- 112.↵
- 113.↵
- 114.↵